24 July 2020 | Author: Jim Lovett
This newsletter was sent via email to those who subscribe to our email updates. Please take a moment to add or confirm your email address with us to receive future updates by completing the short form at https://monarchwatch.org/subscribe
Greetings and welcome to Monarch Watch’s 29th tag season!
As you’ve probably come to expect in the fall, we have a TON of information to share so let’s get to it. This update includes:
1. Monarch Population Status
2. Tagging: then, now and in the future
3. Submitting Tag Data for the 2019 Season
4. Monarch Watch Tagging Kits
5. Tagging Wild and Reared Monarchs
6. Chip in for Monarch Watch in 2020!
7. Grant-Sponsored Free Milkweed Programs
8. Monarch Waystations
9. Collect Milkweed Seed for Monarch Watch
10. Monarch Calendar Project
11. Fall Monarch Watch Events
12. Monarch Rearing, Tagging and Releasing Survey
—————————————————————-
1. Monarch Population Status —by Chip Taylor
—————————————————————-
There are a number of ways to assess the development of the summer monarch breeding population that lend some predictability to the size of the migratory population. One can use mid-summer numbers gleaned from surveys conducted by the North American Butterfly Association or eggs per stem counts tallied by the Monarch Larval Monitoring Project, or simply follow the chatter about the numbers of adult monarchs and immatures reported on various social media sites and email lists like Dplex-L. All have merit and all have limitations.
I prefer to break down the numbers of first sightings north of 40N from 1 May to 14 June as reported to Journey North. I look for two things, the timing of arrival across the longitudes as well as the numbers arriving and I look at how those numbers might have been affected by the numbers of first sightings of returning monarchs from 1 March–30 April. In addition, I ask how the weather might have affected recolonization. To that, I add the forecasts for the mean temperatures for the months of June–August. The result is a prediction each July that is correct more often than it is wrong, but there have been some misses and those are both instructive and humbling.
So, what do these metrics suggest for the coming migration? Actually, the prospects for a reasonably robust migratory population look quite good for the area from central Michigan to 100W in the Dakotas. The numbers from Michigan to the east coast are likely to be lower than for each of the last two years. In terms of the timing of arrivals in the northeast east of Toronto in Canada, and most of northern New England, the recolonization this year was the 6th lowest in the last 21 years. That said, this region has surprised me in the past by producing more monarchs than I expected.
Overall, there should be plenty of monarchs to tag this fall and there is a good likelihood that the numbers this winter in Mexico will be higher than in 2019.
Western Monarchs
The big question for those interested in western monarchs is whether the population will increase from the low numbers each of the last two years. The population in the east bounced back from a low of 0.67 hectares in the winter of 2013–2014 to 4.01 hectares in 2015–2016. That was a remarkable rebound and it’s reasonable that the western population should be able to do the same, right? Maybe and maybe not.
The conditions in the west in terms of the weather extremes, the distribution of milkweeds, etc. are very different from those in the east. Here is what we can say about the west at this point: there is a mix of favorable and not so favorable reports for the west, but generally, there are few reports and that is not a good sign. The good reports involve what appear to be good breeding populations in the vicinity of Prescott, Arizona, and in SE Arizona. Good numbers have also been reported in Salt Lake City and the surrounding area with some reports from Idaho. In contrast, Oregon and Washington have been relatively silent and that is not a good sign. Since a substantial number of the monarchs that arrive at the overwintering colonies along the west coast originate from the Northwest, low production there again this year could mean low numbers during this year’s Thanksgiving Day Counts. Let’s hope things pick up in the west over the next 6 weeks!
—————————————————————-
2. Tagging: then, now and in the future
—————————————————————-
Over the years, thousands of taggers have contributed to our tagging database. It is an enormous record and a veritable gold mine of information about how the migration functions. The record represents over 2 million tagged butterflies and lists where, when and by whom each butterfly was tagged. The sex of each butterfly and whether the butterfly was wild-caught or reared, tagged and released is also recorded. The record also includes over 19,000 recoveries at the overwintering sites. We have spent the last several years closely examining these records and the data have been used as the basis for two publications. The first, “Is the Timing, Pace, and Success of the Monarch Migration Associated With Sun Angle?” was published in December 2019 and the second, “Evaluating the migration mortality hypothesis using monarch tagging data”, should be published soon. Two additional papers are in preparation and more are planned.
Briefly, the tagging data have revealed new information on the origins of monarchs that reach Mexico, the timing and pace of the migration, differences among regions due to recolonization and weather, the impact of drought years and many other factors. None of these insights into the dynamics of the migration and the monarch annual cycle would have been possible without the assistance of all those who have so generously donated their time and data to the Monarch Watch Tagging Database (which will ultimately be transferred to a national archive). Yet, in spite of these successes, we are not done; there is more to learn. The climate is changing and monarch habitats are continuing to decline and for these reasons it is likely that the migration will change as well. Continued tagging should enable us to track these changes, and for that, we hope you will continue to tag, to report your data and to generally support monarch conservation by creating habitats for monarchs or helping others do so.
Thanks, and best of luck this tagging season. Please stay safe. Chip Taylor, Director, Monarch Watch
—————————————————————-
3. Submitting Tag Data for the 2019 Season
—————————————————————-
Thousands of you have already submitted your 2019 season tag data to us by mail or via our online submission form – thank you! We are still receiving data sheets and if you haven’t submitted your data yet it is not too late. Please review the “Submitting Your Tagging Data” information on the tagging program page then send us your data via the Tagging Data Submission Form.
Complete information is available at monarchwatch.org/tagging if you have questions about submitting your data to us and we have conveniently placed a large “Submit Tagging Data Here” button on our homepage at monarchwatch.org that will take you directly to the online form.
There you can upload your data sheets as an Excel or other spreadsheet file (PREFERRED; download a template file from monarchwatch.org/tagging) or a PDF/image file (scan or photo).
If you have any questions about getting your data to us, please feel free to drop Jim a line anytime via JLOVETT@KU.EDU
—————————————————————-
4. Monarch Watch Tagging Kits
—————————————————————-
Monarch tagging is an important tool to help us understand the overall dynamics of the monarch population. Tags for the 2020 fall tagging season are available and we have started shipping them out, ahead of the migration in your area. If you would like to tag monarchs this year, please order your tags soon as they are going fast! Tagging Kits should arrive within 7–10 days but priority will be given to preorders and areas that will experience the migration first.
Monarch Watch Tagging Kits are only shipped to areas east of the Rocky Mountains. Each tagging kit includes a set of specially manufactured monarch butterfly tags (you specify quantity), a data sheet, tagging instructions, and additional monarch / migration information. Tagging Kits for the 2020 season start at only $15 and include your choice of 25, 50, 100, 200, or 500 tags.
Monarch Watch Tagging Kits and other materials (don’t forget a net!) are available via the Monarch Watch Shop online at shop.monarchwatch.org – where each purchase helps support Monarch Watch.
2020 datasheets and instructions are also available online at monarchwatch.org/tagging
Tagging should begin in early to mid-August north of 45N latitude (e.g. Minneapolis), late August at other locations north of 35N (e.g., Oklahoma City, Fort Smith, Memphis, Charlotte) and in September and early October in areas south of 35N latitude. See a map and a table with expected peak migration dates at monarchwatch.org/tagging
—————————————————————-
5. Tagging Wild and Reared Monarchs
—————————————————————-
The following is an abbreviated version of our “Tagging wild and reared monarchs: Best practices” article posted to our Blog last year. The complete text of the article is available via the link below.
Diving into the data has revealed a number of surprises such as the difference between the probability that a reared monarch will reach Mexico and the probability that a wild–tagged monarch will do so. The recovery rate is higher for wild–caught monarchs (0.9% vs 0.5%) and it is the data from the wild–caught butterflies that tell us the most about the migration. Frankly, for some analyses, we have to set the reared monarch data aside. That doesn’t mean it is not valuable, but its uses are limited.
It should be noted that for tagging data purposes, monarchs captured as adult butterflies should be reported as WILD and adult monarchs reared from the egg, larva, or pupa stage should be considered REARED.
TAGGING WILD-CAUGHT MONARCHS
For wild-caught monarchs we need to:
1. increase the number of taggers from western Minnesota and Iowa westward into Nebraska and the Dakotas to give us a more complete understanding of dynamics of the migration;
2. increase the number of wild monarchs that are tagged since these provide the most valuable data; and
3. increase the number of taggers who tag from the beginning of the tagging season in early August until the migration ends. Tagging records for the entire season will help us establish the proportion of the late–season monarchs that reach the overwintering sites. When tagging wild–caught monarchs, many taggers run out of tags well before the season ends. That’s great, but it would help us to know when all tags had been used by indicating this via the online tagging data submission form.
TAGGING REARED MONARCHS
Reared butterflies tend to average smaller than wild migrants. That difference can be reduced significantly if careful attention is given to rearing larvae under the best possible conditions. Large monarchs have the best chance of reaching Mexico, surviving the winter and reproducing in Texas. There are several reasons for this: better glide ratio, better lift with cross or quartering winds, larger fat bodies, more resistance to stress, etc. There are very few small monarchs among those that return in the spring. For those of you who prefer to rear, tag and release, we have a few suggestions:
1. Rear larvae under the most natural conditions possible.
2. Provide an abundance of living or fresh-picked and sanitized foliage to larvae.
3. Provide clean rearing conditions.
4. Plan the rearing so that the newly-emerged monarchs can be tagged early in the migratory season (10 days before to 10 days after the expected date of arrival of the leading edge of the migration in your area).
5. Tag the butterflies once the wings have hardened and release them the day after emergence if possible.
6. When it comes to tagging, tag only the largest and most-fit monarchs (see complete article for some guidelines). Records of tags applied to monarchs that have little chance of reaching Mexico add to the mass of tagging data, but do not help us learn which monarchs reach Mexico – unless the measurements, weight and condition of every monarch tagged and released is recorded. There are a few taggers who keep such detailed records and those data can be very informative. If you collect such data and are willing to share it please contact us; do not add this information to the standard tagging data sheet.
As a final note, this text is not a directive. We are not telling you what to do; rather, we are simply providing suggestions that may lead to more successful rearing and tagging efforts.
The expanded version of this article “Tagging wild and reared monarchs: Best practices” is available at monarchwatch.org/blog/tagging-best-practices
—————————————————————-
6. Chip in for Monarch Watch in 2020!
—————————————————————-
Several years ago, we launched our first “Chip in for Monarch Watch” Fundraising Campaign in honor of our director and founder, Chip Taylor (whose birthday happens to be at the end of August, by the way). This campaign offers a chance for Monarch Watchers, colleagues, friends, and family across the planet to show their support for Chip and the Monarch Watch program he brought to life more than a quarter-century ago. It has provided tremendous support for Monarch Watch over the years, through both monetary contributions and kind words. We encourage you to spend a little time reading through the previous donor comments on the Chip in for Monarch Watch page – the connections that are facilitated by monarchs and Monarch Watch are truly extraordinary.
This year’s Chip in for Monarch Watch campaign is now underway – if you are in a position to offer financial support to Monarch Watch (or know someone who might be), please consider making a fully tax-deductible donation of any amount during our 2020 “Chip in for Monarch Watch” fundraising campaign.
Visit monarchwatch.org/chip for more information or to submit your pledge and tax-deductible donation.
Please be sure to leave your comments, thanks, birthday wishes, etc. for Chip via the Chip in for Monarch Watch Form at form.jotform.com/82141035824146
Thank you for your continued support!
Quick Links:
– Chip in for Monarch Watch: monarchwatch.org/chip
– Chip in for Monarch Watch Form (to leave comments, thanks, birthday wishes, etc.): form.jotform.com/82141035824146
– Donate Now via the KU Endowment: kuendowment.org/monarch
—————————————————————-
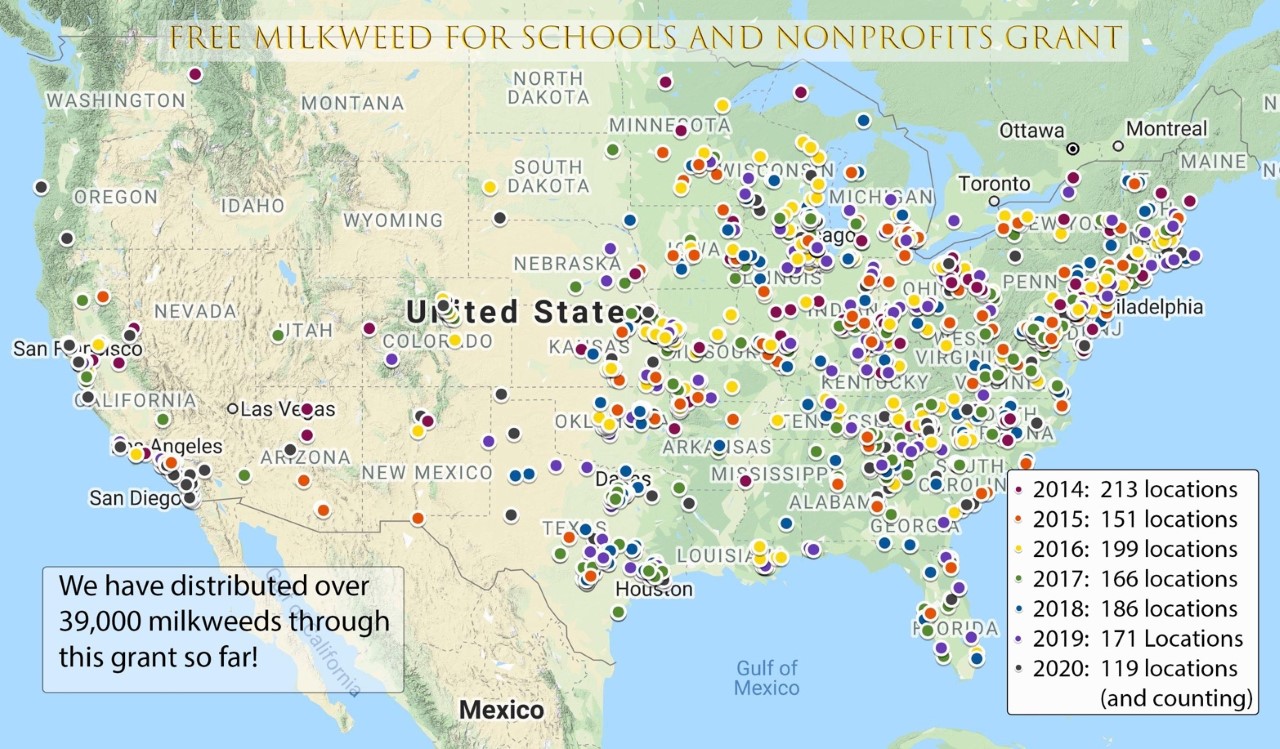
7. Grant-Sponsored Free Milkweed Programs
—————————————————————-
Monarch Watch is still seeking a few more applications for our very popular grant, Free Milkweed for Schools and Nonprofits. If your organization already has a garden and you want to enhance it with milkweeds, this grant is for you! If your organization doesn’t have a garden at all, but would like to start one, this grant is for you! We award these plants to libraries, schools, nature centers, churches, community gardens, scouting and 4H organizations, master gardeners and much more. Help us meet our annual quota of 6,000 plants so that we can reapply for future funding. We are almost there, with less than 2,000 plants to distribute.
The following map shows the distribution of milkweeds since 2014, all funded through the generous support of the Natural Resources Defense Council:
monarchwatch.org/milkweed/free/nonprofit-grant.jpg
Apply now for plants that will be shipped in the fall to the eastern United States. California and Texas applicants will receive plants next spring, when we have a new crop. Awardees will receive one free flat of native milkweed plugs! Sorry, private gardens are not eligible for this grant. Previous grant recipients are not eligible.
For complete details about applying for our Free Milkweeds Grants please visit monarchwatch.org/free-milkweeds
—————————————————————-
8. Monarch Waystations
—————————————————————-
To offset the loss of milkweeds and nectar sources we need to create, conserve, and protect monarch butterfly habitats. You can help by creating “Monarch Waystations” in home gardens, at schools, businesses, parks, zoos, nature centers, along roadsides, and on other unused plots of land. Creating a Monarch Waystation can be as simple as adding milkweeds and nectar sources to existing gardens or maintaining natural habitats with milkweeds. No effort is too small to have a positive impact.
Have you created a habitat for monarchs and other wildlife? If so, help support our conservation efforts by registering your habitat as an official Monarch Waystation today!
monarchwatch.org/waystations
A quick online application will register your site and your habitat will be added to the online registry (mapped location will be approximate for privacy). You will receive a certificate bearing your name and your habitat’s ID that can be used to look up its record. You may also choose to purchase a metal sign to display in your habitat to encourage others to get involved in monarch conservation.
As of 24 July 2020, there have been 29,809 Monarch Waystation habitats registered with Monarch Watch! Texas holds the #1 spot with 2,451 habitats and Illinois (2,320), Michigan (2,233), California (1,909), Ohio (1,529), Virginia (1,379), Florida (1,324), Pennsylvania (1,322), Wisconsin (1,304), and Ontario (978) round out the top ten.
You can view the complete listing and a map of approximate locations via monarchwatch.org/waystations/registry
—————————————————————-
9. Collect Milkweed Seed for Monarch Watch
—————————————————————-
Monarch Watch is seeking donations of milkweed seed collected from wild populations of native milkweeds in order to maintain our milkweed distribution programs. Milkweeds are beginning to put on seed pods and will continue for the next few months, depending upon your location. We welcome seed donations of one ounce or more, which we pass on to our partner nurseries to grow. Through the Milkweed Market and our free milkweed programs, we then distribute milkweed plugs back to the ecoregion from which the seed was collected.
Generally, our current needs include seed from Texas, Arizona, California, the Midwest, and the eastern/southeastern coastal states. Also, incarnata and tuberosa are always needed from east of Ohio, where they are native. We currently have enough seed from the upper Midwest.
Please visit our seed collecting page for further instructions: monarchwatch.org/bring-back-the-monarchs/milkweed/seed-collecting
—————————————————————-
10. Monarch Calendar Project
—————————————————————-
If you are recording your monarch observations for this project and would like to submit your data for the spring (1st) period please do so using the appropriate form, based on your location:
For locations SOUTH of (less than) 35N latitude (March 15–April 30, 2020) please use this form: forms.gle/pBVsLPyJ4xiriuXV8
For locations NORTH of (greater than) 35N latitude (April1–June 20, 2020) please use this form: forms.gle/sfmks7w1cvhD3ghB7
As soon as the fall period ends for all locations (September 25) we will send out links for submission of that data.
Monarch Watch continues to seek the assistance of hundreds of monarch enthusiasts (citizen scientists) in collecting observations of monarchs in their area during specific periods of the spring and fall. If you would like to participate, all you have to do is: 1. REGISTER (just so we know where you are located and how to reach you), 2. RECORD (keep a record of the number of monarch butterflies you observe each day during the time period dictated by your location) and 3. SUBMIT (at the end of the observation period in the fall we will send participants a link to an online form to submit their observation data).
Complete details and a link to the short registration form are available at
monarchwatch.org/calendar
Please note that the fall observation period for those of you in the Southern U.S. runs from August 1–September 25 so you need to start recording your monarch observations! For northern locations (greater than 35N latitude) the fall period runs from July 15–August 20 so recording should be going on now, but it is not too late to join in!
—————————————————————-
11. Fall Monarch Watch Events
—————————————————————-
As you might have guessed, our big in-person public events that are held in September each year in Lawrence, KS (Fall Open House and Tagging Event) have been canceled. However, we hope to be able to provide some alternate activities to engage Monarch Watchers. Stay tuned and stay safe!
—————————————————————-
12. Monarch Rearing, Tagging and Releasing Survey
—————————————————————-
Do you raise monarchs from eggs, caterpillars or chrysalises? If so, help us understand what techniques monarch enthusiasts, such as yourself, are using when rearing monarchs. We have created a simple survey, here: monarchwatch.org/rearing-survey
Feel free to share this with friends or groups that focus on monarch rearing and/or conservation.
Questions? Comments? Please email Angie at monarch@ku.edu
Thank you!
—————————————————————-
About This Monarch Watch List
—————————————————————-
Monarch Watch (https://monarchwatch.org) is a nonprofit education, conservation, and research program affiliated with the Kansas Biological Survey at the University of Kansas. The program strives to provide the public with information about the biology of monarch butterflies, their spectacular migration, and how to use monarchs to further science education in primary and secondary schools. Monarch Watch engages in research on monarch migration biology and monarch population dynamics to better understand how to conserve the monarch migration and also promotes the protection of monarch habitats throughout North America.
We rely on private contributions to support the program and we need your help! Please consider making a tax-deductible donation. Complete details are available at https://monarchwatch.org/donate or you can simply call 785-832-7374 (KU Endowment Association) for more information about giving to Monarch Watch.
If you have any questions about this email or any of our programs, please feel free to contact us anytime.
Thank you for your continued interest and support!
Jim Lovett
Monarch Watch
https://monarchwatch.org
You are receiving this mail because you were subscribed to the Monarch Watch list via monarchwatch.org or shop.monarchwatch.org – if you would rather not receive these periodic email updates from Monarch Watch (or would like to remove an old email address) you may UNSUBSCRIBE via https://monarchwatch.org/unsubscribe
If you would like to receive periodic email updates from Monarch Watch, you may SUBSCRIBE via https://monarchwatch.org/subscribe
This e-mail may be reproduced, printed, or otherwise redistributed as long as it is provided in full and without any modification. Requests to do otherwise must be approved in writing by Monarch Watch.
Filed under Email Updates | Comments Off on Monarch Watch Update July 2020
 In a press release today, the U.S. Fish and Wildlife Service announced its decision with respect to the petition to declare the monarch a “threatened” species under the Endangered Species Act:
In a press release today, the U.S. Fish and Wildlife Service announced its decision with respect to the petition to declare the monarch a “threatened” species under the Endangered Species Act:










 For all of you who are located close (or close enough) to Monarch Watch here in Lawrence, Kansas our Spring Plant Fundraiser is now online at
For all of you who are located close (or close enough) to Monarch Watch here in Lawrence, Kansas our Spring Plant Fundraiser is now online at 





{kind=link}